

MFehi adipose tissue macrophages compensate for tissue iron perturbations in mice
- PMID: 29768045
- PMCID: PMC6171041
- DOI: 10.1152/ajpcell.00103.2018
MFehi adipose tissue macrophages compensate for tissue iron perturbations in mice
Abstract
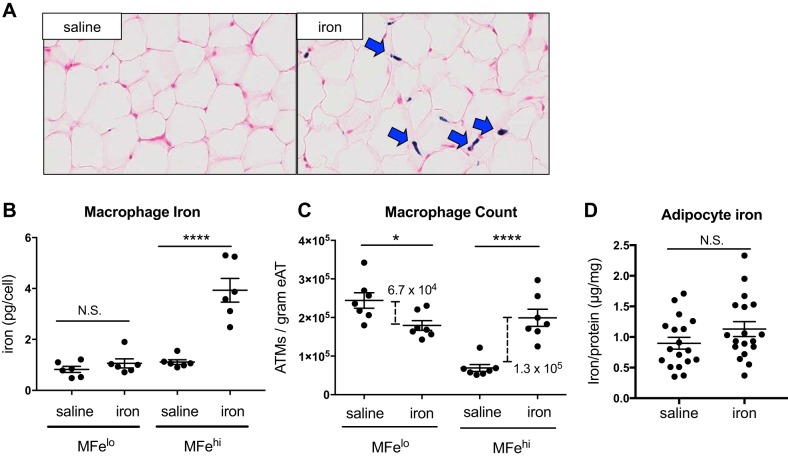
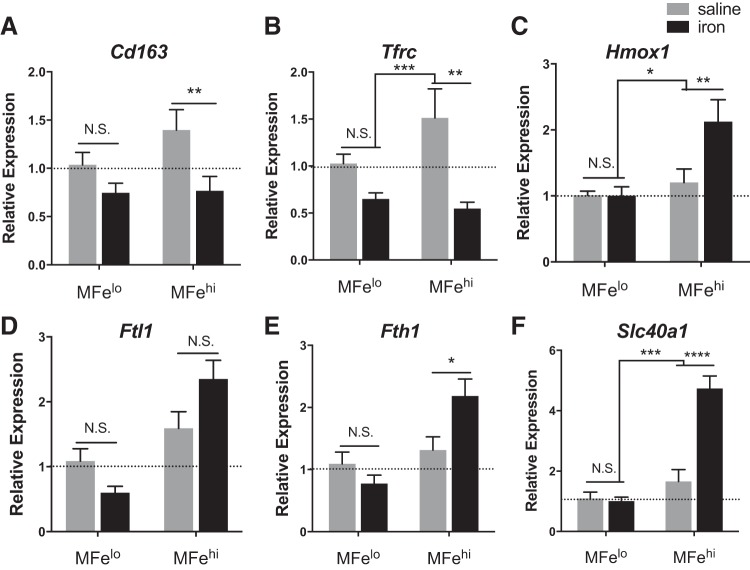
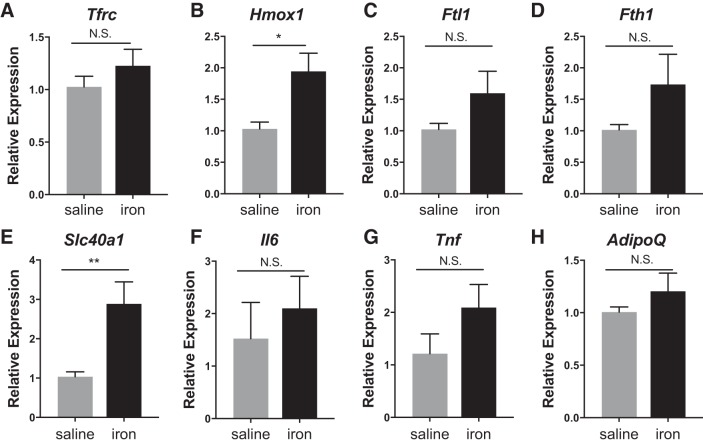
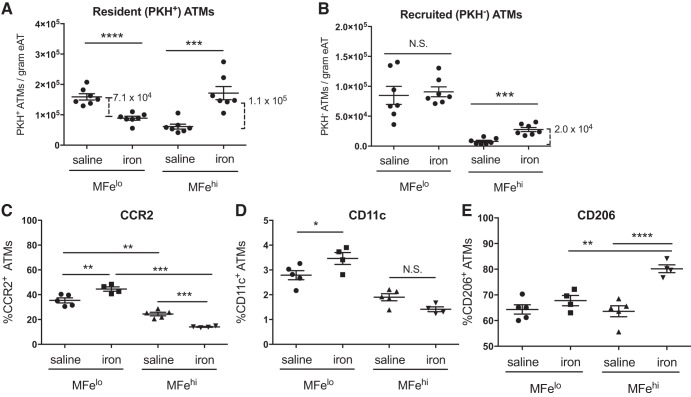
Resident adipose tissue macrophages (ATMs) play multiple roles to maintain tissue homeostasis, such as removing excess free fatty acids and regulation of the extracellular matrix. The phagocytic nature and oxidative resiliency of macrophages not only allows them to function as innate immune cells but also to respond to specific tissue needs, such as iron homeostasis. MFehi ATMs are a subtype of resident ATMs that we recently identified to have twice the intracellular iron content as other ATMs and elevated expression of iron-handling genes. Although studies have demonstrated that iron homeostasis is important for adipocyte health, little is known about how MFehi ATMs may respond to and influence adipose tissue iron availability. Two methodologies were used to address this question: dietary iron supplementation and intraperitoneal iron injection. Upon exposure to high dietary iron, MFehi ATMs accumulated excess iron, whereas the iron content of MFelo ATMs and adipocytes remained unchanged. In this model of chronic iron excess, MFehi ATMs exhibited increased expression of genes involved in iron storage. In the injection model, MFehi ATMs incorporated high levels of iron, and adipocytes were spared iron overload. This acute model of iron overload was associated with increased numbers of MFehi ATMs; 17% could be attributed to monocyte recruitment and 83% to MFelo ATM incorporation into the MFehi pool. The MFehi ATM population maintained its low inflammatory profile and iron-cycling expression profile. These studies expand the field's understanding of ATMs and confirm that they can respond as a tissue iron sink in models of iron overload.
Keywords: adipose; homeostasis; iron; macrophage; polarization.
Figures
References
-
- Bories G, Colin S, Vanhoutte J, Derudas B, Copin C, Fanchon M, Daoudi M, Belloy L, Haulon S, Zawadzki C, Jude B, Staels B, Chinetti-Gbaguidi G. Liver X receptor activation stimulates iron export in human alternative macrophages. Circ Res 113: 1196–1205, 2013. doi: 10.1161/CIRCRESAHA.113.301656. - DOI - PMC - PubMed
-
- Britton L, Jaskowski LA, Bridle K, Secondes E, Wallace D, Santrampurwala N, Reiling J, Miller G, Mangiafico S, Andrikopoulos S, Subramaniam VN, Crawford D. Ferroportin expression in adipocytes does not contribute to iron homeostasis or metabolic responses to a high calorie diet. Cell Mol Gastroenterol Hepatol 5: 319–331, 2018. doi: 10.1016/j.jcmgh.2018.01.005. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous