

Selection of DNA Cleavage Sites by Topoisomerase II Results from Enzyme-Induced Flexibility of DNA
- PMID: 30713098
- PMCID: PMC6474810
- DOI: 10.1016/j.chembiol.2018.12.003
Selection of DNA Cleavage Sites by Topoisomerase II Results from Enzyme-Induced Flexibility of DNA
Abstract
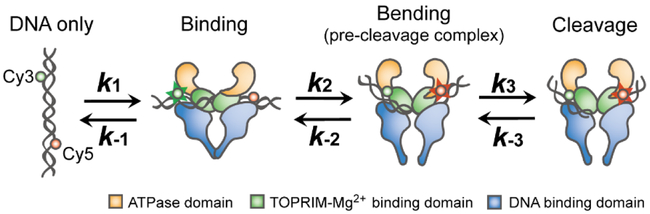
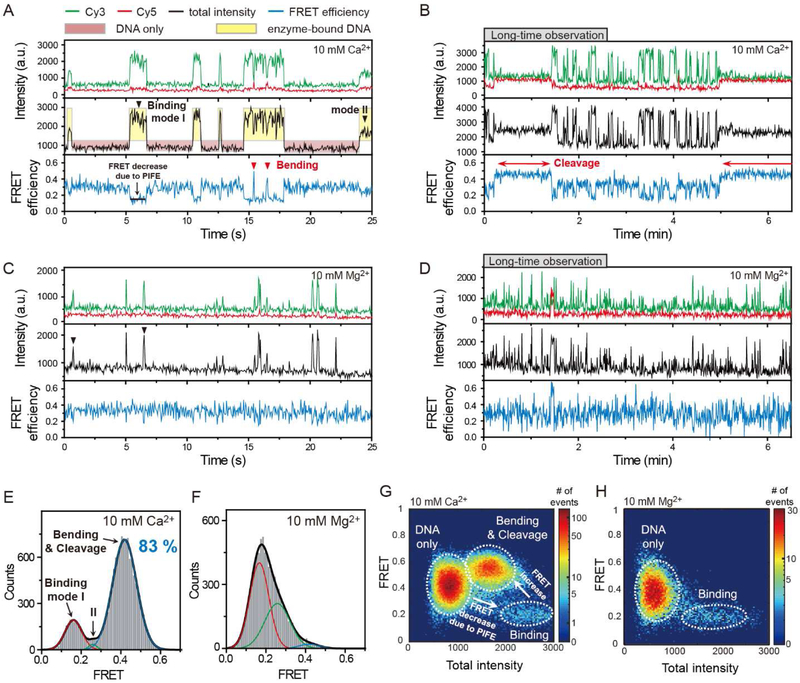
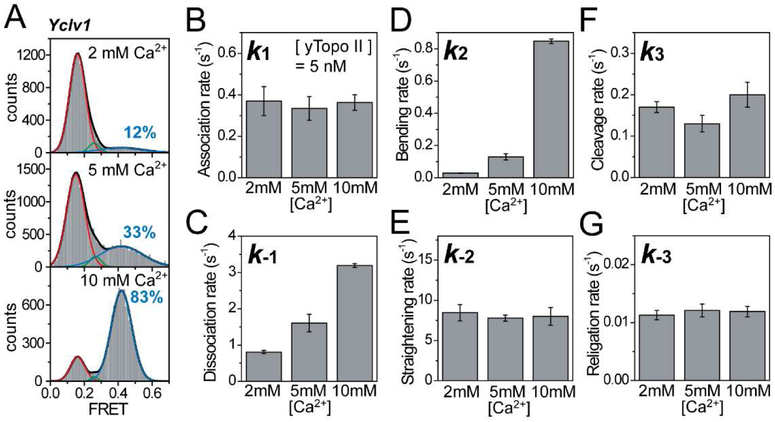
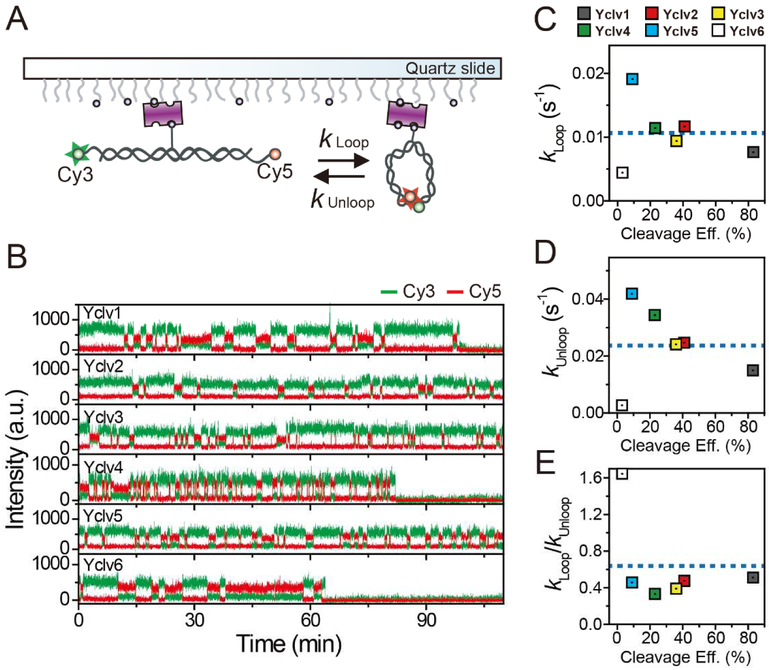
Topoisomerase II cleaves DNA at preferred sequences with different efficiencies; however, the mechanism of cleavage site selection is not known. Here we used single-molecule fluorescence assays that monitor several critical steps of DNA-topoisomerase II interactions, including binding/dissociation, bending/straightening, and cleavage/religation, and reveal that the cleavage site is selected mainly during the bending step. Furthermore, despite the sensitivity of the bending rate to the DNA sequence, it is not an intrinsic property of the DNA itself. Rather, it is determined by protein-DNA interactions.
Keywords: DNA bending; DNA cleavage; DNA flexibility; G-segment selection; sequence preference; single-molecule FRET; topoisomerase II; two-metal-ion mechanism.
Copyright © 2018 Elsevier Ltd. All rights reserved.
Figures


References
-
- Bax BD, Chan PF, Eggleston DS, Fosberry A, Gentry DR, Gorrec F, Giordano I, Hann MM, Hennessy A, Hibbs M, et al. (2010). Type IIA topoisomerase inhibition by a new class of antibacterial agents. Nature 466, 935–940. - PubMed
-
- Berger JM, Gamblin SJ, Harrison SC, and Wang JC (1996). Structure and mechanism of DNA topoisomerase II. Nature 379, 225–232. - PubMed
-
- Bromberg KD, Hendricks C, Burgin AB, and Osheroff N (2002). Human topoisomerase IIα possesses an intrinsic nucleic acid specificity for DNA ligation. Use of 5' covalently activated oligonucleotide substrates to study enzyme mechanism. J Biol Chem 277, 31201–31206. - PubMed
-
- Bromberg KD, and Osheroff N (2001). DNA cleavage and religation by human topoisomerase IIα at high temperature. Biochemistry 40, 8410–8418. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
